
The Case for 2 mSv/day. 2.0.

Another rehash combining past posts and a very difficult read. The goal is to distill the nearly 200 GKN posts down to a basic 5 or 6. This post outlines the fundamentals of radiation damage and repair. You do need to know that radiation dose is measured in two units: grays (Gy) and sieverts (Sv). A gray is the radioactive energy in joules absorbed by a kilogram of tissue. A sievert attempts to adjust the dose in grays for a particle's harmfulness. For most nuclear power plant release purposes, the two units are numerically interchangable.
You can find a properly referenced version at the Flop book website.

Up to 1950, the International Commission on Radiological Protection (ICRP) dose rate limit for the general public was 2 mSv/d. This was called the tolerance dose [rate]. In 1980, Lauriston Taylor, founding chairman of the NCRP, pointed out we have not reliably detected any harm to anybody at this dose rate.

Figure 1. ICRP Dose Limits
However, in 1951, the ICRP changed the recommended limit to 3 mSv/week. This was based on claims of genetic mutations at low doses which turned out to have no foundation.\cite{calabrese-2011} In 1957, the American counterpart of the ICRP, the National Council for Radiation Protection(NCRP), added a limit of 50 mSv/y for nuclear workers and 5 mSv/y for the public. As the NCRP itself acknowledged, this humongous change, a reduction of 30, was not based on any new data.
The changes in the accumulated MPD [Maximum Permissible Dose] are not the result of positive evidence of damage due to use of earlier permissible dose levels but rather are based on the desire to bring the MPD into accord with the trends of scientific opinion.\cite[page 1]{nbs59-1957}
Opinion trends that are not based on data are hardly scientific.
The Green Table shows that this is still the case. To see detectable harm --- get out of the green --- requires dose rates of 20 mSv/d or more, regardless of the cumulative dose. Taylor's 2 mSv/d provides a factor of 10 margin. It's a good rule. But why is this a good rule? To answer that, we must bring in some basic biology.

The concern is DNA damage which if misrepaired can lead to cancer. The DNA in our bodies is constantly being assaulted by Reactive Oxygen Species(ROS). These chemical active molecules such as OH- are the by product of our oxygen based metabolism. About one-billion ROS micro-bombs per day per cell leak from our cell's mitochrondia into the rest of the cell. Roughly 1 in 20 thousand of these molecules chemically damage our DNA. This is the price we pay for an oxygen based metabolism.
DNA damage can take the form of Single Strand Breaks(SSB) and Double Strand Breaks(DSB).1 Table A shows some estimates of the number of SSB's and DSB's each of our cells endure per day from our normal metabolism.

Table A. Metabolic SSB's and DSB's per cell-day
In response to this onslaught, Nature has equipped us with a remarkably accurate DNA repair system. Without this system, we would not be here. SSB's are repaired almost automatically by the clever design of the double helix. The repair uses the intact strand as a template and is essentially error-less. SSB repair takes about 25 minutes.
We are also quite good at repairing isolated DSB's. Molecules called Ku's are always drifting around in the cell waiting for a DSB to happen. When one occurs, they quickly --- in a matter of seconds --- clamp on to the broken ends. The Ku's then call in a large complex of proteins to do the rejoining.
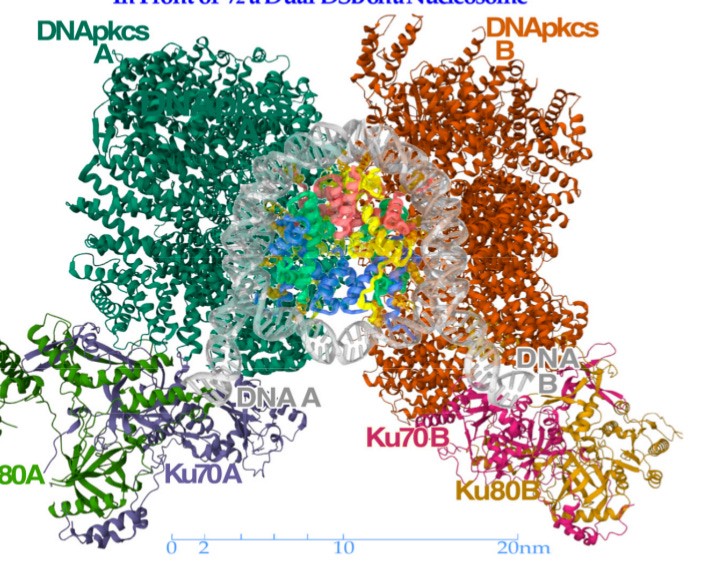
Figure 2. Green and Red are repair RIF's working on translucent DNA with a double DSB. The DSB's are labeled DNA A and DNA B.\cite{brahme-2023}[Figure 9] The Ku's quickly identify and clamp on to the broken ends, and then call in the DNA-Pkcs' to reconnect them.
By tagging the right proteins with fluorescent molecules, these repair clusters, called RIF's (Radiation Induced Foci,) can be viewed, Figure 3. This process, known as an NHEJ repair, does not use a template; but for isolated DSB's, NHEJ almost always works. Even if NHEJ does not get it quite right, some misrepairs can be corrected later in the cell cycle when the cell is preparing for replication. At this point, a template is available and a proof reading process takes place. This template dependent correction is called an HR repair. But this only works for relatively small residual errors.

Figure 3. UCB pictures of cell repair. The bright spots in the three screenshots are clusters of damage sensing and repair proteins, dubbed Radiation Induced Foci (RIF). Berkeley found that the number of RIF's increases less than linearly with dose. At 0.1 Gy, they observed 73 RIF's/Gy. At 1.0 Gy, they saw 28 RIF's/Gy. If a RIF is faced with a single DSB, the repair is almost always correct. If a RIF is faced with more than one DSB, the error rate skyrockets. We expect 25 to 40 DSB's per gray. Do the math. 40 DSB's and 73 RIF's, no problem. 40 DSB's and 28 RIF's, trouble.
Berkeley found that the repair time varied from less than an hour to a day, increasing with dose rate.
The real problem is closely spaced DSB's, which have been dubbed double double strand breaks (DDSB), a rare case of the acronym being better than the name. The RIF's are larger in size than the portions of the DNA they are attempting to repair, Figure 2.\cite{brahme-2023} If the DSB's are too close together, multiple RIF's simply do not have room to form. This would explain why UCB found that the number of RIF's did not rise proportionally with the number of DSB's.\cite{bissell-2011}
When a single RIF is faced with multiple DSB's, it can end up rejoining the wrong ends, creating a possibly viable misrepair.2 A few of the viable mutations will escape our immune system, and a few of those could become cancerous.
If double DSB's are the real problem, then dose rate and repair time becomes all important. The probability that a hit will cause a DDSB is proportional to the inventory of still unrepaired DSB's at the time of the hit.3 To over-simplify, if the repair processes can keep up with the damage, and keep that inventory low, we are OK. If the damage rate is higher than the repair rate, the inventory of unrepaired DSB's will build up, and the probability of a DDSB and a misrepair will grow rapidly.
The Green Table concurs. At very high dose rates, we see significant increases in cancer at cumulative doses as low as 100 mSv. But if the dose rate is below 20 mSv/d, we have been unable to reliably detect any cancer even when the cumulative dose is 100,000 mSv.
Table 2 shows estimates of the DSB's per millisievert of radiation.

If we conservatively assume 10 metabolic DSB's per cell-day, and 0.04 DSB's per millisievert then it would take 250 mSv per day to equal the number of DSB's produced by our metabolism. 250 mSv is about 25,000 times normal background radiation. If normal metabolic damage is equivalent to 250 mSv/d, then any damage associated with 2 mSv/d would almost certainly not be detectable. At the same time, it is not surprising that we start to detect harm at 20 or so mSv/d. At that point, the cell is forced to deal with a substantially higher than normal number of DSB's.
In short, the Table A and Table 2 numbers are consistent with the Green Table, and the pre-1950 tolerance dose rate and its implied repair period of a day.4 2 mSv per day is 700 times higher than the NRC limit of allowed exposure to the public of 1 mSv per year and 3000 times higher than the EPA limit of 0.25 mSv/y, both of which are criminally sold and tragically regarded as harm based numbers.
Since we cannot detect harm at 2 mSv/d, there is no argument for enduring the very real and very large costs of evacuation at this dose rate. At Fukushima, this means no member of the public should have been evacuated, avoiding the nearly immediate killing of 50 frail, elderly people, and the 2300 plus premature deaths associated with the multi-year evacuation. The radiation release would have been a footnote to the massive damage caused by the tsunami, including the loss of three multi-billion dollar reactors. The other three reactors at Fukushima Daichi would have been back on line in a matter of weeks or months. None of the other Japanese nuclear plants would have been affected, avoiding a 30 billion dollar per year increase in electricity costs,\cite{neidell-2019} 2000 to 7000 premature deaths per year due to people being unable to afford heat or cooling,\cite{he-2019}, and a 150 million ton per year increase in CO2 emissions.\cite{kharecha-2019}.
If an overly conservative 2 mSv/d were the evacuation trigger at Chernobyl, only the 50,000 people in the immediate neighborhood of the plant would need to evacuate; and they could have returned to their homes in a matter of weeks. Almost all the harm that was inflicted on 340,000 permanent evacuees would have been avoided. It is telling that Chernobyl Units 1 and 2, which share the same building as the obliterated Chernobyl Unit 4, were back on line in about six months. Most of the plant workers never left.
If our legal and regulatory systems recognized that dose rates of 2 mSv/day and below produce at most negligible harm, nuclear power could be cheaper than coal, as it was in the mid-1960's, when nascent nuclear was just starting down a steep learning curve. A solution to the Gordian Knot of energy poverty and global warming would be within reach.
Some of the damage involves chemical changes other than a break in the strand; but, as long as one side is intact, we call it a Single Strand Break.
A DSB that is not rejoined, can't replicate, and can't cause cancer. Unrepaired cells just die. This is only a problem when there are so many unrepaired cells that we start to see Acute Radiation Syndrome (ARS). The dose rate profiles required to produce clinical ARS have never been inflicted on the public in any nuclear power plant release.
If the repair time is constant, this suggests that the harm will be about quadratic in dose rate. If the repair time increases with dose rate, as Berkeley found, then harm will be something like cubic in dose rate.
This reasoning applies only to the low end, the dose rates that will be experienced by the public in a nuclear plant release. At extremely high doses and dose rates, we see downward curvature in cancer incidence. One possibility is that at such dose rate profiles, the repair process becomes damaged and disrupted. Less repairs means less misrepairs. But that's just a guess at the cause of the high end curvature.
Lauriston Taylor was careful to point out that the tolerance dose [rate] was not a threshold in any absolute sense.
The adoption of tolerance or permissible doses did not depend upon the assumed existence of a threshold. The setting of the tolerance level was on the basis of whether or not effects could be observed, and this has been the common approach to similar problems for all other toxicological agents.\cite{taylor-1971}[page 13]
When my wife applied for the Royal Canadian Mounted Police (RCMP) in 1996, she was moving along the process and in one day in the spring of 1996, as a part of this process, she received about 100 x-rays. They do this in the event an unfortunate situation arises, the police-person perishes and they need help assisting in identifying the body. She asked, "is this many x-rays in one day safe?" She was told "oh yeah, its fine, we've been doing this for years". She also had to get a few dental x-rays for this process and then later in the year, she broke her leg, which ended the hope of becoming an RCMP officer, and had to get a few more x-rays for this.
Also, I understand there is a 10 to 20 msv radiation dose with a whole body CT scan. https://www.cancer.gov/about-cancer/diagnosis-staging/ct-scans-fact-sheet
In Canada, the protection standard is 1msv/year (I assume above background radiation), but of course it varies all over the country.
So I assume occasional instances where SNT is exceeded, even by 5x or more, is OK? Thank you for your work, SNT seems far more rational and practical than LNT.
Note re NRX cleanup.
My father was working at the Chalk River Labs in another building when the accident occurred, so he was involved with the cleanup. He died at age 89 in 2012. More famously Jimmy Carter was also involved at died at age 100.
Clearly 2 lives cruelly cut short by radiations /s
I recall dad mentioning that he went home in Deep River & measured the radioactivity on his clothes which he left on the porch of the house. The next morning he measured the radiation again and found near background. It was almost all very short lived isotopes.